Модель «хищник - жертва» и макроэкономическая модель Гудвина
Рассмотрим биологическую модель «хищник - жертва», в которой один вид является кормом для другого. Эта модель, давно ставшая классической, была построена в первой половине XX в. итальянским математиком В. Вольтерра для объяснения колебаний уловов рыбы в Адриатическом море . В модели предполагается, что число хищников растет до тех пор, пока у них будет достаточно пищи, а увеличение поголовья хищников приводит к уменьшению популяции рыб-жертв. Когда последних становится мало, численность хищников уменьшается. Вследствие этого с некоторого момента начинается рост численности рыб-жертв, который через некоторое время вызывает рост популяции хищников. Цикл замыкается.
Пусть N x (t) и N 2 (t) - численности рыб-жертв и рыб-хищников в момент времени t соответственно. Предположим, что темп прироста численности жертв в условиях отсутствия хищников постоянен, т. е.
где а - положительная постоянная.
Появление хищника должно снизить темп прироста жертв. Будем считать, что это снижение линейно зависит от численности хищника: чем больше хищников, тем меньше темп прироста жертв. Тогда

где т > 0.
Поэтому для динамики численности рыб-жертв получаем:
Составим теперь уравнение, определяющее динамику популяции хищников. Предположим, что их численность в случае отсутствия жертв сокращается (из-за отсутствия корма) с постоянным темпом Ь, т. е.
![]()
Наличие жертв вызывает увеличение темпа прироста хищников. Предположим, что этот прирост линеен, т. е.
![]()
где п> 0.
Тогда для скорости роста рыб-хищников получаем уравнение:
В системе «хищник - жертва» (6.17)-(6.18) снижение скорости роста численности рыб-жертв, вызванное поеданием их хищниками, равно mN x N 2 , т. е. пропорционально числу их встреч с хищником. Увеличение скорости роста численности рыб-хищников, вызванное наличием жертв, равно nN x N 2 , т. е. тоже пропорционально числу встреч жертв и хищников.
Введем безразмерные переменные U = mN 2 /a и V = nN x /b. Динамика переменной U соответствует динамике хищников, а динамика переменной V - динамике жертв. В силу уравнений (6.17) и (6.18) изменение новых переменных определяется системой уравнений:
Предположим, что при t = 0 число особей обоих видов известно, следовательно, известны начальные значения новых переменных?/(0) = U 0 , К(0) = К 0 . Из системы уравнений (6.19) можно найти дифференциальное уравнение для ее фазовых траекторий:

Разделяя переменные этого уравнения, получим:


Рис. 6.10. Построение фазовой траектории ADCBA системы дифференциальных уравнений (6.19)
Отсюда с учетом начальных данных следует:
где константа интегрирования С = b(V Q - InV 0)/a - lnU 0 + U 0 .
На рис. 6.10 показано, как строится линия (6.20) при заданном значении С. Для этого в первой, во второй и в третьей четвертях соответственно строим графики функций х = V - InV, у = (Ь/а)х , у = = InU- U+C.
В силу равенства dx/dV = (V- 1)/У функция х = V- In К, определенная при V > 0, возрастает, если V> 1, и убывает, если V 1. В силу того, что cPx/dV 1 = 1/F 2 > 0, график функции л: = x(V) направлен выпуклостью вниз. Уравнение V= 0 задает вертикальную асимптоту. Наклонных асимптот эта функция не имеет. Поэтому график функции х = х(У) имеет вид кривой, изображенной в первой четверти рис. 6.10.
Аналогичным образом исследуется функция у = InU - U+ С, график которой на рис. 6.10 изображен в третьей четверти.
Если теперь мы разместим на рис. 6.10 во второй четверти график функции у = (Ь/а)х , то в четвертой четверти получим линию, которая связывает переменные U и V. Действительно, взяв точку V t на оси OV, вычисляем с помощью функции х = V - V соответствующее знание х х. После этого, используя функцию у = (Ь/а)х , по полученному значению х { находим у х (вторая четверть на рис. 6.10). Далее с помощью графика функции у = InU - U + С определяем соответствующие значения переменной U (на рис. 6.10 таких значений два - координаты точек М и N). Совокупность всех таких точек (V; U) образует искомую кривую. Из построения следует, что графиком зависимости (6.19) является замкнутая линия, содержащая внутри себя точку Е( 1, 1).
Напомним, что эту кривую мы получили, задав некоторые начальные значения U 0 и V 0 и вычислив по ним константу С. Взяв другие начальные значения, мы получим другую замкнутую линию, не пересекающую первую и также содержащую внутри себя точку Е( 1, 1). Это означает, что семейством траекторий системы (6.19) на фазовой плоскости (V, U) является множество замкнутых непересекающихся линий, концентрирующихся вокруг точки Е( 1, 1), а решения исходной модели U = Щ) и V = V{t) являются функциями, периодическими во времени. При этом максимум функции U = U(t) не попадает на максимум функции V = V(t ) и наоборот, т. е. колебания численности популяций около своих равновесных решений происходят в разных фазах.
На рис. 6.11 приведены четыре траектории системы дифференциальных уравнений (6.19) на фазовой плоскости OUV, отличающиеся начальными условиями. Одна из траекторий равновесная - это точка Е( 1, 1), которой соответствует решение U(t) = 1, V{t) = 1. Точки (U(t), V(t)) на других трех фазовых траекториях по мере увеличения времени смещаются по часовой стрелке.
Чтобы пояснить действие механизма изменения численности двух популяций, рассмотрим траекторию ABCDA на рис. 6.11. Как видим, на участке АВ и хищников, и жертв мало. Поэтому здесь популяция хищников сокращается из-за недостатка корма, а популяция жертв растет. На участке ВС численность жертв достигает высоких значений, что приводит к росту численности хищников. На участке СА хищников много, и это влечет сокращение численности жертв. При этом после прохождения точки D численность жертв уменьшается на столько, что численность популяции начинает уменьшаться. Цикл замыкается.
Модель «хищник - жертва» - пример структурно неустойчивой модели. Здесь малое изменение правой части одного из уравнений может привести к принципиальному изменению ее фазового портрета.

Рис. 6.11.

Рис. 6.12.
Действительно, если в уравнении динамики жертв учесть внутривидовую конкуренцию, то мы получим систему дифференциальных уравнений:
Здесь при т = 0 популяция жертв развивается согласно логическому закону.
При т Ф 0 ненулевое равновесное решение системы (6.21) при некоторых положительных значениях параметра внутривидовой конкуренции И является устойчивым фокусом, и соответствующие траектории «наматываются» на точку равновесия (рис. 6.12). Если же h = 0, то в этом случае особая точка Е( 1, 1) системы (6.19) является центром, и траектории представляют собой замкнутые линии (см. рис. 6.11).
Замечание. Обычно под моделью «хищник - жертва» понимают модель (6.19), фазовые траектории которой замкнуты. Однако модель (6.21) также является моделью «хищник - жертва», поскольку описывает взаимовлияние хищников и жертв.
Одним из первых приложений модели типа «хищник - жертва» в экономике для исследования циклично изменяющихся процессов является макроэкономическая модель Гудвина, в которой используется непрерывный подход к анализу взаимовлияния уровня занятости и ставки заработной платы.
В работе В.-Б. Занга изложен вариант модели Гудвина, в котором производительность труда и предложение труда растут с постоянными темпами прироста, а коэффициент выбытия фондов равен нулю. Эта модель формально приводит к уравнениям модели «хищник - жертва».
Ниже рассматривается модификация этой модели для случая отличного от нуля коэффициента выбытия фондов.
В модели используются следующие обозначения: L - численность трудящихся; w - средняя ставка заработной платы трудящихся; К - основные производственные фонды (капитал); Y - национальный доход; / - инвестиции; С - потребление; р - коэффициент выбытия фондов; N - предложение труда на рынке рабочей силы; т = Y/K - фондоотдача; а = Y/L - производительность труда; у = L/N - коэффициент занятости; х = C/Y - норма потребления в национальном доходе; К - приращение капитала в зависимости от инвестиций.
Выпишем уравнения модели Гудвина:

где а 0 , b, g, п, N 0 , г - положительные числа (параметры).
Уравнения (6.22) - (6.24) выражают следующее. Уравнение (6.22) - обычное уравнение динамики фондов. Уравнение (6.23) отражает рост ставки заработной платы при высоком уровне занятости (ставка заработной платы растет, если предложение труда невелико) и уменьшение ставки заработной платы при высоком уровне безработицы.
Тем самым уравнение (6.23) выражает закон Филлипса в линейной форме . Уравнения (6.24) означают экспоненциальный рост производительности труда и предложения труда. Предположим также, что С = wL, т. е. вся заработная плата тратится на потребление. Теперь можно преобразовать уравнения модели с учетом равенств:

Преобразуем уравнения (6.22)-(6.27). Имеем:
 где
где
где
![]()
Поэтому динамика переменных в модели Гудвина описывается системой дифференциальных уравнений:

которая формально совпадает с уравнениями классической модели «хищник - жертва». Это значит, что в модели Гудвина тоже возникают колебания фазовых переменных. Механизм колебательной динамики заключается здесь в следующем: при малой заработной плате w потребление низкое, инвестиции большие, и это приводит к росту производства и занятости у. Большая занятость у вызывает рост средней заработной платы w, что приводит к росту потребления и снижению инвестиций, падению производства и снижению занятости у.
Ниже гипотеза о зависимости ставки процента от уровня занятости рассмотренной модели используется при моделировании динамики однопродуктовой фирмы. Оказывается, что в этом случае при некоторых дополнительных допущениях модель фирмы обладает свойством цикличности рассмотренной выше модели «хищник - жертва».
- См.: Вольтерра В. Указ, соч.; Ризнииенко Г. Ю., Рубин А. Б. Указ. соч.
- См.: Занг В.-Б. Синергетическая экономика. М., 2000.
- См.: Пу Т. Нелинейная экономическая динамика. Ижевск, 2000; Тихонов А. Н.Математическая модель // Математическая энциклопедия. Т. 3. М., 1982. С. 574, 575.
В модели Колмогорова сделано одно существенное допущение: поскольку предполагается, что при то это означает существование в популяции жертв механизмов, регулирующих их численность даже в отсутствие хищников.
К сожалению, подобная формулировка модели не позволяет ответить на вопрос, вокруг которого в последнее время ведется много споров и о котором мы уже упоминали в начале главы: каким образом популяция хищников может оказывать регулирующее воздействие на популяцию жертв с тем, чтобы вся система была устойчива? Поэтому мы вернемся к модели (2.1), в которой механизмы саморегуляции (например, регулирование при помощи внутривидовой конкуренции) в популяции жертв (впрочем, как и в популяции хищников) отсутствуют; поэтому единственным механизмом регуляции численностей, входящих в сообщество видов, являются трофические взаимоотношения между хищниками и жертвами.
Здесь (так что, в отличие от предыдущей модели, Естественно, что решения (2.1) зависят от конкретного вида трофической функции которая, в свою очередь, определяется характером хищничества, т. е. трофической стратегией хищника и защитной стратегией жертвы. Общими для всех этих функций (см. рис. И) являются следующие свойства:
Система (2.1) имеет одну нетривиальную стационарную точку, координаты которой определяются из уравнений
![]()
при естественном ограничении .
Существует еще одна стационарная точка (0, 0), соответствующая тривиальному равновесию. Нетрудно показать, что эта точка - седло, а сепаратрисами являются координатные оси.
Характеристическое уравнение для точки имеет вид
![]()
Очевидно, что для классической вольтерровской модели .
Поэтому величину f можно рассматривать как меру отклонения рассматриваемой модели от вольтерровской.
![]()
стационарная точка - фокус, и в системе появляются колебания; при выполнении противоположного неравенства - узел, и колебания в системе отсутствуют. Устойчивость этого состояния равновесия определяется условием
т. е. существенным образом зависит от вида трофической функции хищника.
Условие (5.5) можно интерпретировать следующим образом: для устойчивости нетривиального равновесия системы хищник - жертва (и тем самым, для существования этой системы) достаточно, чтобы в окрестности этого состояния относительная доля потребляемых хищником жертв возрастала с возрастанием численности жертв. Действительно, доля жертв (от общей их численности), потребляемых хищником, описывается дифференцируемой функцией условие возрастания которой (положительность производной) выглядит как
![]()
Последнее условие, взятое в точке есть не что иное, как условие (5.5) устойчивости равновесия. При непрерывности оно должно выполняться и в некоторой окрестности точки Таким образом, если численности жертв в этой окрестности, то
Пусть теперь трофическая функция V имеет вид, изображенный на рис. 11, а (характерный для беспозвоночных). Можно показать, что для всех конечных значений (поскольку выпукла вверх)
т. е. при любых величинах стационарной численности жертв неравенство (5.5) не выполняется.
Это означает, что в системе с таким видом трофической функции не существует устойчивого нетривиального равновесия. Возможны несколько исходов: либо численности как жертвы, так и хищника неограниченно возрастают, либо (при прохождении траектории вблизи одной из координатных осей) в силу случайных причин численность жертвы или численность хищника станет равной нулю. При гибели жертвы через некоторое время погибнет и хищник, если же сначала погибает хищник, то численность жертвы начнет экспоненциально возрастать. Третий вариант - возникновение устойчивого предельного цикла - невозможен, что легко доказывается.
В самом деле, выражение

в положительном квадранте всегда положительно, если только имеет вид, изображенный на рис. 11, а. Тогда, согласно критерию Дюлака в этой области отсутствуют замкнутые траектории и устойчивого предельного цикла существовать не может.
Итак, можно сделать вывод: если трофическая функция имеет вид, изображенный на рис. 11, а, то хищник не может быть регулятором, обеспечивающим стабильность популяции жертвы и тем самым устойчивость всей системы в целом. Система может быть устойчива только в том случае, когда в популяции жертвы существуют собственные внутренние регулирующие механизмы, например, внутривидовая конкуренция или эпизоотии. Этот вариант регулирования был уже рассмотрен в §§ 3, 4.
Ранее отмечалось, что такой тип трофической функции характерен для хищников-насекомых, «жертвами» которых также обычно являются насекомые. С другой стороны, наблюдения за динамикой многих природных сообществ типа «хищник - жертва», включающих в себя виды насекомых, показывают, что для них характерны колебания очень большой амплитуды и весьма специфического вида.
Обычно после более или менее постепенного роста численности (который может происходить либо монотонно, либо в виде колебаний с возрастающей амплитудой) происходит ее резкое падение (рис. 14), и затем картина повторяется. По-видимому, такой характер динамики численностей видов насекомых может быть объяснен неустойчивостью этой системы при малых и средних значениях численности и действием мощных внутрипопуляционных регуляторов численности при больших ее значениях.

Рис. 14. Динамика численности популяции австралийской листоблошки Cardiaspina albitextura, кормящейся на эвкалиптах. (Из статьи: Clark L. R. The population dynamics of Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, № 3, p. 362-380.)
Если же в систему «хищник - жертва» входят виды, способные к достаточно сложному поведению (например, хищники способны к обучению или жертвы способны находить убежище), то в такой системе возможно существование устойчивого нетривиального равновесия. Доказывается это утверждение весьма просто.
В самом деле, трофическая функция тогда должна иметь вид, изображенной на рис. 11, в. Точка на этом графике - точка касания прямой, проведенной из начала координат, графика трофической функции Очевидно, что в этой точке функция имеет максимум. Также легко показывается, что для всех выполняется условие (5.5). Следовательно, нетривиальное равновесие, в котором численность жертв меньше будет асимптотически устойчиво
Однако мы ничего не можем сказать о том, сколь велика область устойчивости этого равновесия. Например, если существует неустойчивый предельный цикл, то эта область должна лежать внутри цикла. Или другой вариант: нетривиальное равновесие (5.2) неустойчиво, но существует устойчивый предельный цикл; в этом случае также можно говорить о стабильности системы хищник - жертва. Поскольку выражение (5.7) при выборе трофической функции типа рис. 11, в может менять знак при изменении при , то критерий Дюлака здесь не работает и вопрос о существовании предельных циклов остается открытым.

КОМПЬЮТЕРНАЯ МОДЕЛЬ «ХИЩНИК-ЖЕРТВА»
Казачков Игорь Алексеевич 1 , Гусева Елена Николаевна 2
1 Магнитогорский государственный технический университет им. Г.И. Носова, институт строительства, архитектуры и искусства, студент 5 курса
2 Магнитогорский государственный технический университет им. Г.И. Носова, институт энергетики и автоматизированный систем, кандидат педагогических наук, доцент кафедры бизнес-информатики и информационных технологий
Аннотация
Данная статья посвящена обзору компьютерной модели «хищник-жертва». Проведенное исследование позволяет утверждать, что экологическое моделирование играет огромную роль в исследовании окружающей среды. Данная проблематика имеет многогранный характер.
COMPUTER MODEL «PREDATOR-VICTIM»
Kazatchkov Igor Alekseevich 1 , Guseva Elena Nikolaevna 2
1 Nosov Magnitogorsk State Technical University, Civil Engineering, Architecture and Arts Institute, student of the 5th course
2 Nosov Magnitogorsk State Technical University, Power Engineering and Automated Systems Institute, PhD in Pedagogical Science, Associate Professor of the Business Computer Science and Information Technologies Department
Abstract
This article provides an overview of the computer model "predator-victim". The study suggests that environmental simulation plays a huge role in the study of the environment. This problem is multifaceted.
Для исследования окружающей нас среды используют экологическое моделирование. Математические модели используют в тех случаях, когда нет естественной среды и нет естественных объектов, она помогает сделать прогноз влияния разных факторов на исследуемый объект. Данный метод берет на себя функции проверки, построения и интерпретацию полученных результатов. На основе таких форм экологическое моделирование занимается оценкой изменений, окружающей нас среды.
В настоящий момент подобные формы используется для изучения окружающей нас среды, а когда требуется изучить какую-либо из ее областей, то применяют математическое моделирование. Данная модель дает возможность спрогнозировать влияние тех или иных факторов на объект изучения. В свое время был предложен тип «хищник – жертва» такими учеными как: Т. Мальтусом (Malthus 1798, Мальтус 1905), Ферхюльстом (Verhulst 1838), Пирлом (Pearl 1927, 1930), а также А. Лотки (Lotka 1925, 1927) и В. Вольтерры (Volterra 1926).Эти модели воспроизводят периодический колебательный режим, возникающий в результате межвидовых взаимодействий в природе.
Одним из основных методов познания является моделировка. Помимо того, что в нем можно спрогнозировать изменения, происходящие в окружающей среде, к тому же помогает найти оптимальный способ решения проблемы. Уже давно в экологии используют математические модели, для того чтобы установить закономерности, тенденции развития популяций, помогают выделить суть наблюдений. Макет может служить образцом поведения, объекта.
При воссоздании объектов в математической биологии используются прогнозирования различных систем, предусматриваются специальные индивидуальности биосистем: внутренне строение особи, условия жизнеобеспечения, постоянство экологических систем, благодаря которым сберегается жизнедеятельность систем.
Появление компьютерного моделирования значительно раздвинуло рубеж способностей исследования. Возникло вероятность многосторонней реализации трудных форм, не допускающих аналитического изучения, появились новейшие направления, а еще имитационное моделирование.
Рассмотрим, что же такое объект моделирования. «Объектом является замкнутая среда обитания, где происходит взаимодействие двух биологических популяций: хищников и жертв. Процесс роста, вымирания и размножения происходит
непосредственно на поверхности среды обитания. Питание жертв происходит за счет тех ресурсов, которые присутствуют в данной среде, а питание хищников происходит за счет жертв. При этом питательные ресурсы могут быть как возобновляемые, так и не возобновляемые.
В 1931 году Вито Вольтеррой были выведены следующие законы отношения хищник-жертва.
Закон периодического цикла – процесс уничтожения жертвы хищником нередко приводит к периодическим колебаниям численности популяций обоих видов, зависящим только от скорости роста плотоядных и растительноядных, и от исходного соотношения их численности.
Закон сохранения средних величин – средняя численность каждого вида постоянна, независимо от начального уровня, при условии, что специфические скорости увеличения численности популяций, а также эффективность хищничества постоянны.
Закон нарушения средних величин – при сокращении обоих видов пропорционально их числу, средняя численность популяции жертвы растет, а хищников – падает.
Модель хищник-жертва – это особая взаимосвязь хищника с жертвой, в результате которой выигрывают оба. Выживают наиболее здоровые и приспособленные особи к условиям среды обитания, т.е. все это происходит благодаря естественному отбору. В той среде где нет возможности для размножения, хищник рано или поздно уничтожит популяцию жертвы, в последствии чего вымрет и сам» .
На земле существует множество живых организмов, которые при благоприятных условиях увеличивают численность сородичей до огромных масштабов. Такая способность называется: биотический потенциал вида, т.е. увеличение численности вида за определенный промежуток времени. Каждый вид имеет свой биотический потенциал, к примеру крупные виды организмов за год могут возрасти всего в 1,1 раза, в свою очередь организмы более мелких видов, таких как рачки и т.д. могут увеличить свой вид до 1030 раз, ну а бактерии еще в большем количестве. В любом из этих случаев популяция будет расти в геометрической прогрессии.
Экспоненциальным ростом численности называется геометрическая прогрессия роста численности популяции. Такую способность можно наблюдать в лаборатории у бактерий, дрожжей. В не лабораторных условиях экспоненциальный рост возможно увидеть на примере саранчи или же на примере других видов насекомых. Такой рост численности вида можно наблюдать в тех местах где у него практически нет врагов, а продуктов питания более чем достаточно. В конце концов увеличение вида, после того как численность возросла в течении непродолжительного времени, рост популяции начинал снижаться.
Рассмотрим компьютерную модель размножения млекопитающих на примере модели Лотки-Вольтерры. Пусть на некоторой территории обитают два вида животных: олени и волки. Математическая модель изменения численности популяций в модели
Лотки-Вольтерры:
Начальное число жертв - xn, число хищников - yn.
Параметры модели:
P1– вероятность встречи с хищником,
P2– коэффициент роста хищников за счет жертв,
d – коэффициент смертности хищников,
a – коэффициент прироста численности жертв.
В учебной задаче были заданы такие значения: численность оленей равнялось 500, численности волков равна 10, коэффициент прироста оленей равен 0,02, коэффициент прироста численности волков равен 0,1, вероятность встречи с хищником 0,0026, коэффициент роста хищников за счет жертв 0,000056. Данные рассчитаны на 203 года.
Исследуем влияние коэффициент прироста жертв на развитие двух популяций, остальные параметры оставим без изменений.
На схеме 1 наблюдается увеличение численности жертвы и затем, с некоторым опозданием наблюдается прирост хищников. Затем хищники выбивают жертв, число жертв резко падает и вслед за ним уменьшается число хищников (рис. 1).

Рисунок 1. Численность популяций при низкой рождаемости у жертв
Проанализируем изменение модели, увеличив коэффициент рождаемости жертвы а=0,06. На схеме 2 мы видим циклический колебательный процесс, приводящий к увеличению численности обоих популяций со временем (рис. 2).

Рисунок 2.Численность популяций при средней рождаемости у жертв
Рассмотрим как изменится динамика популяций при высоком значении коэффициента рождаемости жертвы а=1,13. На рис. 3 наблюдается резкое увеличение численности обеих популяций с последующим вымиранием, как жертвы, так и хищника. Это происходит за счет того, что численность популяции жертв увеличилось до такого количества, что стали заканчиваться ресурсы, вследствие чего происходит вымирание жертвы. Вымирание хищников происходит из-за того, что сократилось количество жертв и у хищников закончились ресурсы для существования.
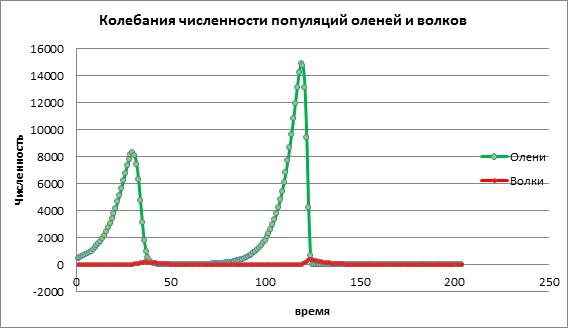
Рисунок 3.Численность популяций при высокой рождаемости у жертв
Исходя из анализа данных компьютерного эксперимента, можно сделать выводы о том, что компьютерное моделирование позволяет нам прогнозировать численность популяций, изучать влияние различных факторов на популяционную динамику. В приведенном примере мы исследовали модель «хищник-жертва», влияние коэффициента рождаемости жертв на численность оленей и волков. Небольшой прирост популяции жертв приводит к небольшому увеличению жертв, которую через некоторый период уничтожают хищники. Умеренный прирост популяции жертв приводит к увеличению численности обеих популяций. Высокий прирост популяции жертв приводит сначала к быстрому росту популяции жертв, это влияет на увеличение роста хищников, но затем расплодившиеся хищники быстро уничтожают популяцию оленей. В итоге оба вида вымирают.
система РА88, которая одновременно предсказывает вероятность более чем 100 фармакологических эффектов и механизмов действия вещества на основе его структурной формулы. Эффективность применения этого подхода к планированию скрининга составляет около 800%, а точность компьютерного прогноза на 300% превосходит предсказание экспертов.
Итак, одним из конструктивных инструментов получения новых знаний и решений в медицине является метод математического моделирования. Процесс математизации медицины – частое проявление взаимопроникновения научных знаний, повышающее эффективность лечебно-профилактической работы.
4. Математическая модель «хищники-жертвы»
Впервые в биологии математическую модель периодического изменения числа антагонистических видов животных предложил итальянский математик В. Вольтерра с сотрудниками. Модель, предложенная Вольтерра, явилась развитием идеи, намеченной в 1924 году А. Лоттки в книге "Элементы физической биологии". Поэтому эта классическая математическая модель известна как модель "Лоттки-Вольтерра".
Хотя в природе отношения антагонистических видов более сложные, чем в модели, тем не менее они являются хорошей учебной моделью, на которой можно изучать основные идеи математического моделирования.
Итак, задача : в некотором экологически замкнутом районе живут два вида животных (например, рыси и зайцы). Зайцы (жертвы) питаются растительной пищей, имеющейся всегда в достаточном количестве (в рамках данной модели не учитывается ограниченность ресурсов растительной пищи). Рыси (хищники) могут питаться только зайцами. Необходимо определить, как будет меняться численность жертв и хищников с течением времени в такой экологической системе. Если популяция жертв увеличивается, вероятность встреч хищников с жертвами возрастает, и, соответственно, после некоторой временной задержки, растет популяция хищников. Эта достаточно простая модель вполне адекватно описывает взаимодействие между реальными популяциями хищников и жертв в природе.
Теперь приступим к составлению дифференциальных уравнений. Обо-
значим число жертв через N, а число хищников через M. Числа N и M являются функциями времени t . В нашей модели учтем следующие факторы:
а) естественное размножение жертв; б) естественная гибель жертв;
в) уничтожение жертв за счет поедания их хищниками; г) естественное вымирание хищников;
д) увеличение числа хищников за счет размножения при наличии пищи.
Так как речь идет о математической модели, то задачей является получение уравнений, в которые входили бы все намеченные факторы и которые описывали бы динамику, то есть изменение числа хищников и жертв со временем.
Пусть за некоторое время t количество жертв и хищников изменится на ∆N и ∆M. Изменение числа жертв ∆N за время ∆t определяется, во-первых, увеличением в результате естественного размножения (которое пропорционально имеющемуся количеству жертв):
где В – коэффициент пропорциональности, характеризующий скорость естественного вымирания жертв.
В основе вывода уравнения, описывающего уменьшение числа жертв изза поедания их хищниками, лежит идея о том, что чем чаще происходит их встреча, тем быстрее уменьшается число жертв. Ясно также, что частота встреч хищников с жертвой пропорционально и числу жертв и числу хищников, то
Поделив левую и правую часть уравнения (4) на ∆t и перейдя к пределу при ∆t→0 , получим дифференциальное уравнение первого порядка:
Для того, чтобы решить это уравнение, нужно знать, как меняется число хищников (М ) со временем. Изменение числа хищников (∆М ) определяется увеличением из-за естественного размножения при наличии достаточного количества пищи (М 1 = Q∙N∙M∙∆t ) и уменьшением из-за естественного вымирания хищников (M 2 = - P∙M∙∆t ):
M = Q∙N∙M∙∆t - P∙M∙∆t |
Из уравнения (6) можно получить дифференциальное уравнение:
Дифференциальные уравнения (5) и (7) представляют собой математическую модель "хищники-жертвы". Достаточно определить значения коэффици-

ентов A, B, C, Q, P и математическую модель можно использовать для решения поставленной задачи.
Проверка и корректировка математической модели. В данной лабора-
торной работе предлагается кроме просчета наиболее полной математической модели (уравнения 5 и 7), исследовать более простые, в которых что-либо не учитывается.
Рассмотрев пять уровней сложности математической модели, можно "почувствовать" этап проверки и корректировки модели.
1-ый уровень – в модели учтено для "жертв" только их естественное размножение, "хищники" отсутствуют;
2-ой уровень – в модели учтено для "жертв" их естественное вымирание, "хищники" отсутствуют;
3-ий уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, "хищники" отсутствуют;
4-ый уровень – в модели учтены для "жертв" их естественное размножение
и вымирание, а также поедание "хищниками", но число "хищников" остается неизменным;
5-ый уровень – в модели учтены все обсуждаемые факторы.
Итак, имеем следующую систему дифференциальных уравнений:
где М – число "хищников"; N – число "жертв";
t – текущее время;
A – скорость размножения "жертв"; C – частота встреч "хищники-жертвы"; B – скорость вымирания "жертв";
Q – размножение "хищников";
P – вымирание "хищников".
1-ый уровень: М = 0, В = 0; 2-ой уровень: М = 0, А = 0; 3-ий уровень: М = 0; 4-ый уровень: Q = 0, Р = 0;
5-ый уровень: полная система уравнений.
Подставляя значения коэффициентов в каждый уровень, будем получать разные решения, например:
Для 3-его уровня значение коэффициента М=0 , тогда

решая уравнение получим
Аналогично для 1-го и 2-го уровней. Что касается 4-го и 5-го уровней, то здесь необходимо решать систему уравнений методом Рунге-Кутта. В результате получим решение математических моделей данных уровней.
II. РАБОТА СТУДЕНТОВ ВО ВРЕМЯ ПРАКТИЧЕСКОГО ЗАНЯТИЯ
Задание 1 . Устно-речевой контроль и коррекция усвоения теоретического материала занятия. Сдача допуска к занятию.
Задание 2 . Выполнение лабораторной работы, обсуждение полученных результатов, оформление конспекта.
Выполнение работы
1. С рабочего стола компьютера вызвать программу "Лаб. №6", щелкнув по соответствующему ярлыку два раза левой клавишей "мыши".
2. Щелкнуть дважды левой клавишей "мыши" по ярлыку "PREDATOR".
3. Выбрать ярлык "PRED" и повторить вызов программы левой клавишей "мыши" (щелкнув дважды).
4. После титульной заставки нажать "ENTER".
5. Моделирование начинать с 1-го уровня.
6. Ввести год, с которого будет проводиться анализ модели: например, 2000
7. Выбрать временные интервалы, например, в течение 40 лет, через 1 год (затем через 4 года).
2-ой уровень: B = 0.05; N0 = 200;
3-ий уровень: A = 0.02; B = 0.05; N = 200;
4-ый уровень: A = 0.01; B = 0.002; C = 0.01; N0 = 200; M = 40; 5-ый уровень: A = 1; B = 0.5; C = 0.02; Q = 0.002; P = 0.3; N0 = 200;
9. Подготовить письменный отчет по работе, который должен содержать уравнения, графики, результаты расчета характеристик модели, выводы по проделанный работе.
Задание 3. Контроль конечного уровня знаний:
а) устно-речевой отчет за выполненную лабораторную работу; б) решение ситуационных задач; в) компьютерное тестирование.
Задание 4. Задание на следующее занятие: раздел и тема занятия, согласование тем реферативных докладов (объем доклада 2-3 стр., регламент 5-7 мин.).
Еще в 20-х гг. А. Лотка (Lotka), а несколько позднее независимо от него В. Вольтерра предложили математические модели, описывающие сопряженные колебания численности популяций хищника и жертвы. Рассмотрим самый простой вариант модели Лотки-Вольтерра. В основе модели лежит ряд допущений:
1) популяция жертв в отсутствие хищника растет экспоненциально,
2) пресс хищников тормозит этот рост,
3) смертность жертв пропорциональна частоте встреч хищника и жертвы (или иначе, пропорциональна произведению плотностей их популяций);
4) рождаемость хищника зависит от интенсивности потребления жертв.
Мгновенная скорость изменения численности популяции жертв может быть выражена уравнением
dN ж /dt = r 1 N ж - p 1 N ж N х,
где r 1 - удельная мгновенная скорость популяционного роста жертвы, p 1 - константа, связывающая смертность жертв с плотностью хищника, a N ж и N х - плотности соответственно жертвы и хищника.
Мгновенная скорость роста популяции хищника в этой модели принимается равной разности рождаемости и постоянной смертности:
dN х /dt = p 2 N ж N х – d 2 N х,
где p 2 - константа, связывающая рождаемость в популяции хищника с плотностью жертв, a d 2 - удельная смертность хищника.
Согласно приведенным уравнениям каждая из взаимодействующих популяций в своем увеличении ограничена только другой популяцией, т.е. рост числа жертв лимитируется прессом хищников, а рост числа хищников - недостаточным количеством жертв. Никакого самоограничения популяций не предполагается. Считается, например, что пищи для жертвы всегда достаточно. Также не предполагается и выхода популяции жертв из-под контроля хищника, хотя на самом деле такое бывает достаточно часто.
Несмотря на всю условность модели Лотки-Вольтерра, она заслуживает внимания уже хотя бы потому, что показывает, как даже такая идеализированная система взаимодействия двух популяций может порождать достаточно сложную динамику их численности. Решение системы этих уравнений позволяет сформулировать условия поддержания постоянной (равновесной) численности каждого из видoв. Популяция жертв сохраняет постоянную численность, если плотность хищника равна r 1 /p 1, а для того чтобы постоянство сохраняла популяция хищника, плотность жертв должна быть равна d 2 /p 2 . Если на графике отложить по оси абсцисс плотность жертв N ж , а по оси ординат - плотность хищника N х , то изоклины, показывающие условие постоянства хищника и жертвы, будут представлять собой две прямые, перпендикулярные друг другу и координатным осям (рис. 6,а). При этом предполагается, что ниже определенной (равной d 2 /p 2) плотности жертв плотность хищника всегда будет уменьшаться, а выше - всегда увеличиваться. Соответственно и плотность жертвы возрастает, если плотность хищника ниже значения, равного r 1 /p 1 , и уменьшается, если, она выше этого значения. Точка пересечения изоклин соответствует условию постоянства численности хищника и жертвы, а другие точки на плоскости этого графика совершают движение по замкнутым траекториям, отражая, таким образом, регулярные колебания численности хищника и жертвы (рис. 6,б). Размах колебаний определяется начальным соотношением плотностей хищника и жертвы. Чем ближе оно к точке пересечения изоклин, тем меньше окружность, описываемая векторами, и соответственно меньше амплитуда колебаний.
Рис. 6. Графическое выражение модели Лотки-Вольтера для системы хищник-жертва.
Одна из первых попыток получения колебаний численности хищника и жертвы в лабораторных экспериментах принадлежала Г.Ф. Гаузе (Gause). Объектами этих экспериментов были инфузория парамеция (Paramecium caudatum ) и хищная инфузория дидиниум (Didinium nasutum ). Пищей для парамеции служила регулярно вносимая в среду взвесь бактерий, а дидиниум питался только парамециями. Данная система оказалась крайне неустойчивой: пресс хищника по мере увеличения его численности приводил к полному истреблению жертв, после чего вымирала и популяция самого хищника. Усложняя опыты, Гаузе устраивал убежище для жертвы, внося в пробирки с инфузориями немного стеклянной ваты. Среди нитей ваты могли свободно перемещаться парамеции, но не могли дидиниумы. В таком варианте опыта дидиниум съедал всех парамеций, плавающих в свободной от ваты части пробирки, и вымирал, а популяция парамеции затем восстанавливалась за счет размножения особей, уцелевших в убежище. Некоторого подобия колебаний численности хищника и жертвы Гаузе удалось добиться только в том случае, когда он время от времени вносил в культуру и жертву и хищника, имитируя таким образом иммиграцию.
Через 40 лет после работы Гаузе его опыты были повторены Л. Лакинбиялом (Luckinbill), использовавшим в качестве жертвы инфузорию Paramecium aurelia , а в качестве хищника того же Didinium nasutum . Лакинбиллу удалось получить несколько циклов колебаний численности этих популяций, но только в том случае, когда плотность парамеций была лимитирована нехваткой пищи (бактерий), а в культуральную жидкость добавляли метилцеллюлозу - вещество, снижающее скорость движения как хищника, так и жертвы и потому уменьшающее частоту их возможных встреч. Оказалось также, что добиться колебаний хищника и жертвы легче, если увеличить объем экспериментального сосуда, хотя условие пищевого лимитирования жертвы и в этом случае обязательно. Если же к системе сосуществующих в колебательном режиме хищника и жертвы добавляли избыточную пищу, то ответом был, быстрый рост численности жертвы, за которым следовало возрастание численности хищника, приводящее в свою очередь к полному истреблению популяции жертвы.
Модели Лотки и Вольтерры послужили толчком для разработки ряда других более реалистичных моделей системы хищник-жертва. В частности, довольно простая графическая модель, анализирующая соотношение разных изоклин жертвы хищника, была предложена М. Розенцвейгом и Р. Мак-Артуров (Rosenzweig, MacArthur). Согласно этим авторам, стационарная (= постоянная) численность жертвы в координатных осях плотности хищника и жертвы может быть представлена в виде выпуклой изоклины (рис. 7,а). Одна точка пересечения изоклины сосью плотности жертвы соответствует минимальной допустимой плотности жертвы (нижеее популяция подвержена очень большому риску вымирания хотя бы из-за малой частоты встреч самцов и самок), а другая - максимальной, определяемой количеством имеющейся пищи или поведенческими особенностями самой жертвы. Подчеркнем, что речь идет пока о минимальной и максимальной плотностях в отсутствие хищника. При появлении хищника и увеличении его численности минимальная допустимая плотность жертвы, очевидно, должна быть выше, а максимальная - ниже. Каждому значению плотности жертвы должна соответствовать некоторая плотность хищника, при которой достигается постоянство популяции жертвы. Геометрическое место таких точек и есть изоклина жертвы в координатах плотности хищника и жертвы. Векторы, показывающие направление изменения плотности жертвы (ориентированные горизонтально), имеют разную направленность по разные стороны от изоклины (рис. 7,а).

Рис. 7. Изоклины стационарных популяций жертвы (а) и хищника (б).
Для хищника в тех же координатах также построена изоклина, отвечающая стационарному состоянию его популяции. Векторы, показывающие направление изменения численности хищника, ориентированы вверх или вниз в зависимости от того, по какую сторону от изоклины они находятся. Форма изоклины хищника, показанная на рис. 7,б. определяется, во-первых, наличием некоторой минимальной плотности жертвы, достаточной для поддержания популяции хищника (при более низкой плотности жертвы хищник не может увеличивать свою численность), а во-вторых, наличием некоторой максимальной плотности самого хищника, при превышении которой численность будет снижаться независимо от обилия жертв.

Рис. 8. Возникновение колебательных режимов в системе хищник-жертва в зависимости от расположения изоклин хищника и жертвы.
При совмещении изоклин жертвы и хищника на одном, графике возможны три различных варианта (рис. 8). Если изоклина хищника пересекает изоклину жертвы в том месте, где она уже снижается (при высокой плотности жертв), векторы, показывающие изменение численности хищника и жертвы, образуют траекторию, закручивающуюся во внутрь, что соответствует затухающим колебаниям численности жертвы и хищника (рис. 8,а). В том случае, когда изоклина хищника пересекает изоклину жертвы в ее восходящей части (т.е. в области низких значений плотности жертв), векторы образуют раскручивающуюся траекторию, а колебания численности хищника и жертвы происходят соответственно с возрастающей амплитудой (рис. 8,б). Если же изоклина хищника пересекает изоклину жертвы в области ее вершины, то векторы образуют замкнутый круг, а колебания численности жертвы и хищника характеризуются стабильной амплитудой и периодом (рис. 8,в).
Иными словами, затухающие колебания соответствуют ситуации, при которой хищник ощутимо воздействует на популяцию жертв, достигнувшую только очень высокой плотности (близкой к предельной), а колебания возрастающей амплитуды возникают, тогда, когда хищник способен быстро увеличивать свою численность даже при невысокой плотности жертв и таким образом быстро ее уничтожить. В других вариантах своей модели Poзенцвейг и Мак-Артур показали, что стабилизировать колебания хищник-жертва можно, введя «убежище», т.е. предположив, что в области низкой плотности жертв существует область, где численность жертвы растет независимо от количества имеющихся хищников.
Стремление сделать модели более реалистичными путем их усложнения проявилось в работах не только теоретиков, но и экспериментаторов. В частности, интересные результаты были получены Хаффейкером (Huffaker), показавшим возможность сосуществования хищника и жертвы в колебательном режиме на примере мелкого растительноядного клеща Eotetranychus sexmaculatus и нападающего на него хищного клеща Typhlodromus occidentalis . В качестве пищи для растительноядного клеща использовали апельсины, помещенные на подносы с лунками (вроде тех, что используются для хранения и перевозки яиц). В первоначальном варианте на одном подносе было 40 лунок, причем в некоторых из них находились апельсины (частично очищенные от кожуры), а в других - резиновые мячики. Оба вида клещей размножаются партеногенетически очень быстро, и поэтому характер их популяционной динамики можно выявить за сравнительно короткий срок. Поместив на поднос 20 самок растительноядного клеща Хаффейкер наблюдал быстрый рост его популяции, которая стабилизировалась на уровне 5-8 тыс. особей (в расчете на один апельсин). Если к растущей популяции жертвы добавляли несколько особей хищника, то популяция последнего быстро увеличивала свою численность и вымирала, когда все жертвы оказывались съеденными.
Увеличив размер подноса до 120 лунок, в которых отдельные апельсины были случайно разбросаны среди множества резиновых мячиков, Хаффейкеру удалось продлить сосуществование хищника и жертвы. Важную роль во взаимодействии хищника и жертвы, как выяснилось, играет соотношение скоростей их расселения. Хаффейкер предположил, что, облегчив передвижение жертвы и затруднив передвижение хищника, можно увеличить время их сосуществования. Для этого на подносе из 120 лунок среди резиновых мячиков располагали случайным образом 6 апельсинов, причем вокруг лунок с апельсинами были устроены преграды из вазелина, препятствовавшие расселению хищника, а для облегчения расселения жертвы на подносе были укреплены деревянные колышки, служившие своего рода «взлетными площадками» для растительноядных клещей (дело в том, что этот вид выпускает тонкие нити и с помощью их может парить в воздухе, распространяясь по ветру). В таком усложненном местообитании хищник и жертва сосуществовали в течение 8 месяцев, продемонстрировав три полных цикла колебаний численности. Наиболее важные условия этого сосуществования следующие: гетерогенность местообитания (в смысле наличия в ней пригодных и непригодных для обитания жертвы участков), а также возможность миграции жертвы и хищника (с сохранением некоторого преимущества жертвы в скорости этого процесса). Иными словами, хищник может полностью истребить то или иное локальное скопление жертв, но часть особей жертвы успеет мигрировать и дать начало другим локальным скоплениям. До новых локальных скоплений хищник рано или поздно тоже доберется, но тем временем жертва успеет расселиться в другие места (в том числе и в те, где она обитала раньше, но потом была истреблена).
Нечто подобное тому, что наблюдал Хаффейкер в эксперименте, встречается и в природных условиях. Так, например, бабочка кактусовая огневка (Cactoblastis cactorum ), завезенная в Австралию, значительно снизила численность кактуса опунции, но не уничтожила его полностью именно потому, что кактус успевает расселиться немного быстрее. В тех местах, где опунция истребляется полностью, перестает встречаться и огневка. Поэтому, когда через некоторое время сюда вновь проникает опунция, то в течение определенного периода она может произрастать без риска быть уничтоженной огневкой. Со временем, однако, огневка снова здесь появляется и, быстро размножаясь, уничтожает опунцию.
Говоря о колебаниях хищник-жертва, нельзя не упомянуть и о циклических изменениях численности зайца и рыси в Канаде, прослеженных по материалам статистики заготовок пушнины компанией Гудзон-Бэй с конца XVIII вплоть до начала XX в. Этот пример нередко рассматривался как классическая иллюстрация колебаний хищник-жертва, хотя на самом деле мы видим только следование роста численности популяции хищника (рыси) за ростом численности жертвы (зайца). Что же касается снижения численности зайцев после каждого подъема, то оно не могло объясняться только возросшим прессом хищников, а было связано с другими факторами, по-видимому, прежде всего нехваткой корма в зимний период. К такому выводу пришел, в частности, М. Джилпин (Gilpin), пытавшийся проверить, могут ли быть описаны эти данные классической моделью Лотки-Вольтерры. Результаты проверки показали, что удовлетворительного соответствия модели нет, но как ни странно, оно становилось лучше, если хищника и жертву меняли местами, т.е. трактовали рысь как «жертву», а зайца - как «хищника». Подобная ситуация нашла свое отражение и в шутливом названии статьи («Едят ли зайцы рысей?»), по сути своей очень серьезной и опубликованной в серьезном научном журнале.